 |
| since2004― |
|
  |


Pet4You.jpのエコ活動

「GIO」と「ECO」のコラボレーション

ベアドッグの育成でお世話になっています

環境との調和をめざすリゾート
オススメ!
|
 |
|
ヒグマ間に働く力学と準テリトリー理論
ヒグマの成長とは生き抜いていくための警戒心を獲得することともいえ、特にオスは成獣に至るまでに各種学習・類推によって警戒心を獲得し、年齢増加とともに孤立性が自律的に加わるのが普通である。
ヒグマにはテリトリー(縄張り)は存在しない。これは数頭のヒグマが行動圏をクロスさせて年間を過ごす事実から従来の生物学的には端的に言えるが、年間のテリトリアルなエリアが存在していないからといってヒグマ相互間に力学が働いていないということではない。ヒグマを捉え対策をおこなうにあたって重要なのは、生態学に加えヒグマ相互間の力学である。
ヒグマ間に働く力学を考えるとき、空間(三次元)に加え時間軸を加味しなければならない。つまり、GPS発信器等で2頭のヒグマの行動圏を把握でき、その行動圏がクロスしていた場合、ある日時に2頭のヒグマがどういう位置関係にあったかが重要で、その状態がどれだけの時間維持され、またどのように変化したかも加味する必要がある。GPS発信器等の遠隔的な調査方法は有益ではあるが、ある個体の位置情報は得られても、発信器をつけていない別個体との位置関係やそこに働いている力が現れない。つまり、ヒグマの社会学的なつながりが隠される盲点がある。そのため、ヒグマの社会学的見地の研究では従来型の現場を歩き回る調査方法が必要不可欠である。
様々なヒグマの観察から、時限的(期限的・期間限定的)にできるあるクマのテリトリアルな空間を「準テリトリー」と私は規定しているが、準テリトリーはシカの死骸・フキの群生地など食物の特異点によってできあがることが多く、一定時間で消滅する性質のものである。ただし、アユのテリトリーで時間軸を加味した場合、準テリトリー的な働き方が一般で、なおかつ個体密度が一定レベルを超えるとテリトリーが消滅することも知られている。これに似た準テリトリー消滅がヒグマにも存在し、その条件として「誘因度が高く豊富な食物」がある。
この時限的で流動的な性質ゆえ、世界各国でヒグマのテリトリーがあるように見えたり、無いように見えたり自在に変わって意見もばらついてきたと考えられる。
ホームレンジはヒグマの存在範囲ではなく通常存在範囲の外周で囲まれた空間であるが、実質的に外周をクローズアップするメリットは少ない。準テリトリーの理論は、ホームレンジをのっぺりとした空間として捉えるのではなく、準テリトリーと移動・徘徊ルートの集合として捉えるところからはじめる。移動・徘徊には比較的無目的な移動、食物を明確にめざした移動から交尾期のメス追尾まで含まれる。
もし仮に、ヒグマ同様私の首にGPS発信器を装着して一年間追跡したとしよう。ウロウロしている範囲はだいたい季節ごとに決まっているが、ときどき旭川や札幌へも行くし、ごく希に実家の東京やフォーラムのために中部地方や中国地方にまで足を運ぶ。この時、年間行動圏はかなり広大な外周を持っているとデータ上はなるが、じゃあ東京へ行く途中の仙台や群馬県に何か意味があるかというと、ない。そして、その年に限って訪れた中国地方をクローズアップして「行動圏が非常に広い」と評価する意味合いが薄いと私自身は考えている。少なくとも、仮にあるクマが遠くに移動した場合、その目的・原動力が何なのかという部分と、そのクマにとってどれだけ意味が重いことなのかを加味して補正をしながら行動範囲を考えたいのである。
さて、準テリトリー理論の概念図が下図である。
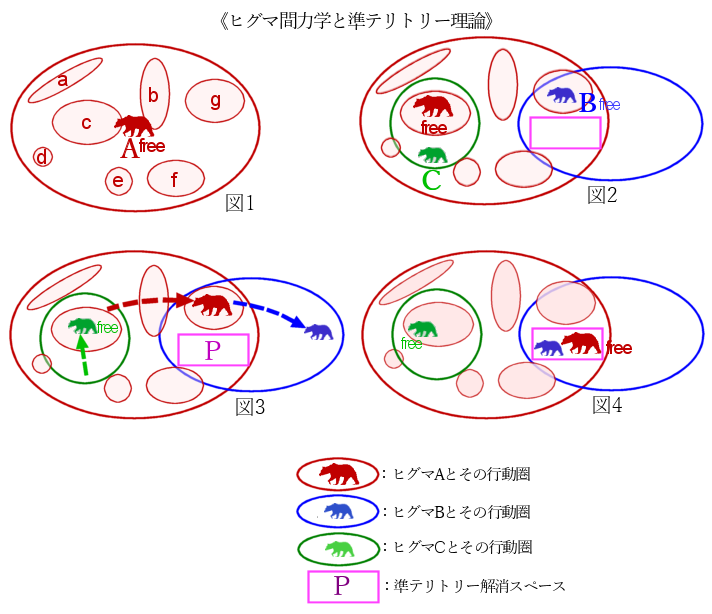
図1
まず、あるヒグマAの行動圏を描いたのが図1だが、その中に特異点となるえさ場や季節によりよく活動している空間があり、その中で特に準テリトリーをa~gで表した。「free」と書いたのは、ほかのクマの動向を気にせず、自分の意志で行きたい場所へ行って好きに活動できる状態を意味する。上図で、ヒグマAはオス成獣で、周辺のクマより優位個体として設定した。
ヒグマ社会の中で最も優位な個体はどのクマの動向も気にせず行動できるが、種を越えた社会まで想定すれば、唯一ヒトの動向を気にして巧みに避けながら行動することになる。
図2
実際はヒグマAだけがこのエリアに存在しているわけでなく、行動圏をクロスさせながら複数頭のいろいろなクマが活動しているだろう。ここではA・B・C3頭のヒグマが活動していたと仮定する。図1のモデルにヒグマB/Cの行動圏を重ねてみる。ヒグマBはAの行動圏の右側で行動圏をクロスさせているが、ヒグマCは若い個体かメス熊で、行動圏がヒグマAの行動圏の中に収まっている。行動圏がクロスしているので、生物学の定義からすれば「ヒグマにはテリトリーはない」と単純に結論できるが、仮にヒグマにとって有利な空間があっても、それぞれが一定のばらつきを持って通常活動しているため、ヒグマ同士に遠ざかる力学が働いていることは、その事実だけからも示唆されている。
この図では、優位個体Aの移動・動向によってB・Cの移動が起きうる。優位なオス成獣Aがヒグマ力学の作用点(トリガー)になっているため、まずヒグマAを考える。ヒグマAが準テリトリーcで活動しているため、ヒグマCはその場所を回避して、十分距離をとって自らの行動圏の別エリアで活動している。この場合、ヒグマAがヒグマBを退けるのではなく、ヒグマAの存在を感知したヒグマBが自ずとヒグマAを避ける、いわば忌避力学が働いている。一方、ヒグマBは自らの行動圏の外でヒグマAが活動しているためfreeな状態となり、自由気ままに自らの行動圏の一等地で活動しているだろう。
図3
準テリトリーCがフキの群生地ならば、ヒグマAはその空間を一定期間占有する形で食べ漁ると別のフキ群生地にに移動することも多い。仮にヒグマAが準テリトリーgに移動したとすると、気ままに暮らしていたヒグマBはヒグマAを気にした暮らしに移行し、やはり悶着を回避するため自らの行動圏内で移動してヒグマAの活動と距離をとる。一方、ヒグマCはほどなくヒグマAが自らの行動圏から出ていったことを感知し、ヒグマAの準テリトリーc付近にも姿を現し、ヒグマAが食べ残したフキを食べることもある。
図4
ある季節、サーモンが海から遡上してきた、あるいは、デントコーンが食べ頃の実をつけはじめた、となると、その場所(図中のP)には準テリトリーは大なり小なりできにくく、複数のヒグマの狭い共有空間ができあがる。そのできにくさは、サーモンの遡上河川周辺で大きく(遡上数にもよる)、デントコーン農地の場合は相対的に小さい。つまり、準テリトリーのできにくさも連続的に程度がある。
説明するためのモデルではなく実際のヒグマ生息地では、20頭もの個体が行動圏をクロスさせて活動しているが、その中心部付近に食物の特異点となる大規模なえさ場があり、ある時期になるとそこを中心にできあがるほとんどのヒグマの共有空間が準テリトリーのできないスペースである。この特異空間をダイニングスペース(Dining
Space)、それをつくる原動力となっている食の特異点をダイニングポイント(Dining Point)と呼ぶことにする。このスペースでは、誘因度の高い食物と狭い空間の共有のせいでヒグマ同士の突発的な(刹那的な)争いが起こる率が高くなるが、それを回避するための工夫も様々おこなわれる。ダイニングポイント脇での順番待ち、ダイニングスペース内でのアピール(背こすり・マーキング糞など)、そして時間を違えたダイニングポイントへの移動などは、ダイニングスペース外より頻繁に、また正確・律儀におこなわれる。
、
逆に、広大なヒグマ生息地からダイニングポイント・ダイニングスペースを見てみよう。他愛のない落書きのような上図は、色を違えて10頭のヒグマの行動圏を模式的に描いた図だ。大きな楕円、だいたいオス成獣の行動圏。中央付近に幾つもある小さな楕円が若グマもしくはメス熊の行動圏である。行動圏の重なりを見ていくと、中央部にどのクマも踏み入る空間があることに気がつく。これが食餌可能な誘因度の高い食物が豊富に存在するダイニングポイントを有したダイニングスペースで、代表的なケースとしてサーモンが遡上する河川の小滝などサーモンが溜まる場所が挙げられる。観光スポットとして整備されているマクニール(アラスカ州)が有名だが、アラスカ・カナダのサーモン遡上河川には大なり小なりマクニール同様ヒグマが集結するポイントが複数箇所あり、そこでは突発的ないざこざやサーモン捕獲場所の順番待ちはあるものの、複数のヒグマの共有空間として機能する。
 北海道では、サーモンの遡上はヒトによって河口部で阻まれこの写真のような光景はなかなか見られないが、再三触れているデントコーン農地というのがこれに類似する特異点を形成している。ヒグマが比較的健全に暮らす生息地に隣接してクマに対して無防備に植えられた大量のデントコーンは、サーモンほど誘引力はないものの、ほとんど同様の状況をつくることが丸瀬布の観察でわかってきている。 北海道では、サーモンの遡上はヒトによって河口部で阻まれこの写真のような光景はなかなか見られないが、再三触れているデントコーン農地というのがこれに類似する特異点を形成している。ヒグマが比較的健全に暮らす生息地に隣接してクマに対して無防備に植えられた大量のデントコーンは、サーモンほど誘引力はないものの、ほとんど同様の状況をつくることが丸瀬布の観察でわかってきている。
直感的に理解できると思うが一応整理しておくと、この特異点・特異空間「ダイニングポイントとスペース」において、ヒグマの行動圏配置にのみ着眼し幾つかの事実を条件として与えると、帰結が簡単に出てくる。
1。若グマ・メス熊は行動圏が狭く、お互いに空間を共有することができる。
2。オス成獣は行動圏が広く、狭い空間でを共有することは苦手である。
帰結:ダイニングスペース周辺においてメス熊・若グマの生息密度が年間を通して高くなり、ダイニングポイントができる時期には、遠征組のオス熊がそれに加わりさらにヒグマの高密度地帯になる。
実際には、メス熊が多いエリアがヒグマ生産エリアになることから、上述の高密度化が速いスピードで起こり過剰になっていくことや、逆にデントコーン農地の場合、ヒトの活動状況によって大型オス成獣がヒトの活動が多い場所を忌避することから、すべてのクマの共有空間ができにくいなど、幾つか修正して考えるべき事があるとともに、例えばメスの高密度エリアになるとすれば、繁殖期には多くのオスがこのエリアに到来する、あるいはその影響で親離れが半ば強制的に(※)促されたり、親離れ直後の若グマがヒトの活動圏に接近してトラブルを起こしやすいなど、派生的に起こる現象もある。
現実的に、丸瀬布「いこいの森」周辺のデントコーン農地周辺では、オス成獣の数が少ない点を除き、ほとんどマクニールと同じ現象が起きている。あるポイントに仕掛けたトレイルカメラ一台に、二週間ほどで10頭以上のヒグマが写ってくるのは、ここのデントコーン農地が準テリトリー解消スペースになっているからなのだ。
※5月後半~7月にかけての交尾期における仔熊を連れたメスの行動パタンは、通常、オスを感知して遠ざかるように移動するか、逆にオスを追うように風下からついていくか、その二つだが、交尾目的でそのエリアに到来するオスの数が一定レベルを超えると、1歳の子を連れた母グマの多くはオスを避けるのではなく、仔熊を親離れさせてオスの交尾を受け入れる戦略を持ち出す。オスの到来数が増える原因としてメスへの性比偏重があるため、結果的にそのエリアの親離れ時期として1歳半前後タイプが多くなり、つまり、メス熊の子育てローテーションの周期が短くなり、ヒグマの生産効率が非常に高いエリアとして機能するようになる。
ちょっと休憩:サーモンの森
私が学生時代から活動をおこなっていたアラスカのYentnaRiver上流部の森では、自宅の裏山にレッドサーモンが累々と遡る山上湖があって、その遡上時期になると、もともとその周辺に暮らしていたヒグマ(Brownbear)に加え、どこからともなく多くのヒグマが集まりヒグマ高密度地帯を形成した。クマのみならず、オオカミ・クズリ・キツネなどほとんどの肉食獣やカケスに始まる鳥類のサーモンへの依存度は高く、レッドサーモンの到来とともにそれはそれは賑やかな森になった。マクニールのクマは観光用に長い時間をかけてヒトに反応しないよう学習させて、なおかつそこを訪れる人のコントロールも精度高くおこなわれているが、Yentnaの森のクマはヒトへの馴化ができていないので、より正確にヒグマのことを知り緻密にクマ対策をおこなって暮らすことを余儀なくされた。そこで観察し推測したことが現在の丸瀬布「いこいの森」周辺ではほとんどそのまま起きており、30年経った今になって、当時私はYentnaの森のヒグマのダイニングスペースに暮らしていたと気づかされた。
北海道のヒトは、観光エリアや人里の中にわざわざ大規模なダイニングポイントをつくり、ヒグマを山のあちこちから効果的に呼び寄せてそこで暮らす。知らぬ顔で観光客も呼ぶ。随分酔狂なことをするものだと感心させられた。 |
では、基本となるヒグマの力学とはどのような力学か。
オオカミのようなテリトリアルなほ乳類の力学は原則的に排他的である。マーキング・ハウリング・パトロールなどによってテリトリーの恒常的かつときにシステマチックな管理をおこなう。すべての個体が比較的正確に認知した方法でテリトリーの誇示をおこないつつ、外部からの侵入個体・侵入パック(群れ)を積極的に排除する手法を持っており、いわば「排他の力学」である。それに対しヒグマの力学は原始的・消極的で、いわば遠慮・敬遠を引き出す「忌避の力学」である。ヒグマの特にオス成獣による樹印・背こすりの少なくとも一部は、誇示ではなくアピールの意味合いが強いと考えられ、そのアピールエリアに他個体が侵入してもオオカミのような排他性・攻撃性は生じないことも各種観察でわかってきた。ヒグマには、単なるいざこざ・喧嘩に相当する争いが発生することがあるが、これは刹那的な気分によるものであってテリトリアルな性質由来ではない。
なお、このヒグマの社会に一定のバランスをもたらす忌避力学を引きだしているヒグマの性質・能力に、先述した類推能力が働いている。
ヒトとヒグマの関係学―――忌避力学とテリトリーの拡張理論
また、私自身が実践し模索してきたヒトとヒグマの共生の方法論は、この忌避力学にヒトを混入させて、いわば優位なオス成獣のごとく各ヒグマに認識させ、ヒトへの忌避・敬遠の心理を引き出すことで、時限的ヒトの活動(ヒグマ生息地における伐採・釣りなど)が準テリトリーとして機能し、また、恒常的ヒトの活動域(人里)がその延長としてテリトリアルな空間として機能するようヒトとヒグマの関係性を構築する方向である。この点、オオカミ・ヒトはテリトリアルでヒグマはそれにあたらない。
テリトリーの概念は、そもそも同種の動物個体間・パック間に定義され当てはめられてきたものだが、同種に限定する必然性が無く、異種間の関係性に拡張することが可能である。
このヒグマの忌避力学をヒグマの社会学モデルに結びつけるために、図2のような(ヒトを含めた)忌避の強さ(忌避度数)をそれぞれ考える。この数値は警戒心に概ね比例するが、警戒心と好奇心・誘引力のトータルとして表現されている。
例)オス熊からヒトへの忌避の強さを10とすると、メス熊からオス熊へは11、若グマ同士は相互に1の忌避が働いている、などと読む。
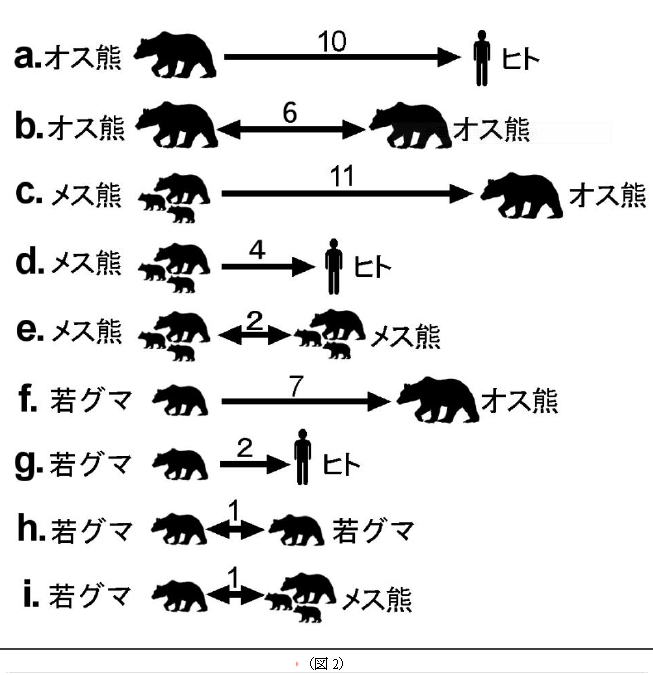
上図でメス熊若グマのオス熊に対する忌避度が高く、それぞれ「11と7」であるが、この高い忌避が原動力となってメス熊・若グマはオスの活動から遠ざかるように行動圏を定める。
また、メス熊・若グマのヒトへの忌避度は「4と2」で低いが、忌避力学的な考えをすれば、「メス熊・若グマはオス熊よりヒトの活動に接近して活動する」と帰結できる。さらに、オス熊のヒトに対する忌避度が強く(10)、アラスカの研究者から聞いた表現では「メス熊はヒトを利用してオス熊を避ける」となるし、メス熊・若グマのヒトに対する忌避度が下がっていけば、より居心地の良い安全な場所として人里周りが認知されうるため、メス熊・若グマはヒトや人里周辺が安全であると積極的に確認しがちである、とも言える。もし仮に、そこに大規模なダイニングポイントがあればメス熊・若グマにとっては願ってもない好適な生活場所となる。
 オス成獣とて闇雲に周辺の同族を攻撃したりするわけではない。しかし、誘因度・執着度の高い食物が絡んだケースで若グマが軽率に行動していれば、何かの拍子にオス熊の逆鱗に触れ攻撃を受けることがあり、右写真のようにたった一撃で致命傷に近い怪我を負うこともなくはない。この若グマが生き延びたかどうかはわからないが、通常、親離れしたあとのまだ学習途上の若グマはここまで重症にならずにオス成獣を警戒・忌避し、距離を十分とって活動するようになる。 オス成獣とて闇雲に周辺の同族を攻撃したりするわけではない。しかし、誘因度・執着度の高い食物が絡んだケースで若グマが軽率に行動していれば、何かの拍子にオス熊の逆鱗に触れ攻撃を受けることがあり、右写真のようにたった一撃で致命傷に近い怪我を負うこともなくはない。この若グマが生き延びたかどうかはわからないが、通常、親離れしたあとのまだ学習途上の若グマはここまで重症にならずにオス成獣を警戒・忌避し、距離を十分とって活動するようになる。
ベアドッグを用いた追い払いなどの威圧・威嚇を「若グマの忌避教育」と位置づけそう呼んでいるが、上図の忌避力学的にいうと、それぞれ低いメス熊・若グマからのヒトに対する忌避度を積極的・意図的に上げてやり、結果的にヒトの活動場所からそれらのクマの暮らしを遠ざけることが一つの目的となっている。忌避度でいえば、ちょうど若グマからオス熊に対する度数あたり、つまり「7」程度の忌避度を引き出すことを目標としている。
ただ、ダイニングポイント・ダイニングスペースの解消は、それらができる条件のうち「食餌可能な」という部分を消してやる以外に不可能で、クマ用の電気柵が普及する以外、延々人里がダイニングスペースに含まれる状況は続くだろう。
※注1)インテリジェンスフローにしたがいヒグマの個体差が大きく、また、成獣においても体験によって変化するので、この忌避度数は変動値であり、また、だいたい正しい数値を用いて相対的に説明してはいるが、あくまで定性的なモデルと捉えて欲しい。残念ながら、定量化を伴った忌避力学の理論はパラメータが複雑すぎて見出せていない。
※注2)電気柵や箱罠では、ヒグマの行動改善や不注意な個体のアトランダムな捕殺排除は一定レベルでできても意識改善がおこなわれない。そのため、最終的にヒトによるヒグマの管理あるいはヒトとヒグマの共生は、ヒグマの忌避力学にヒトを加え、各ヒグマにヒトを意識させながら距離をとらせる・忌避・遠慮させる・敬遠させるという関係学的なコントロールの方法になるだろう。
ちょっと休憩:テリトリーの目的とメンテナンス
「テリトリー・なわばり」というと、なんとなく業突張りでケチな感じも受けるが、本来的にその個体もしくはパック(群れ・集団)が他個体・多集団とトラブルを起こさないためのて合理的な方法論である。それぞれが自由気ままに縦横無尽に動き回り、いつバッタリと出合い争いになるかわからないより、きっちり棲み分けをおこなってしまったほうがその種にとって安定的で好都合な場合にテリトリーは形成され、また効率的に機能する。
テリトリーはその種の性質に加え特に食物を中心とした生活環境によって広さ・強度が可変で、例えば、アユのテリトリーでは個体密度が一定レベルを超えると、テリトリーの消失が起こる。こういう現象は、はじめに定型的なルールや決まりがあるのではなく、合理性を考えてシンプルに理解することができる場合が多い。つまり、アユの場合なら、アユはそれぞれ河床の石に付いたコケをできるだけ多く食みたいためにテリトリーをつくってある範囲のコケを独占したいわけだが、のべつ幕なしに他個体が侵入してきてそれを追い払っていると、肝心の「コケを食べる」という行為自体ができなくなる。そこで、進入個体が一定レベルを超えると、いちいち体当たりをして追い払うのをやめ、無視してひたすら食べるようになる。
アユのテリトリーでわかるように、テリトリーという概念そのものに空間的・時間的要素が規定されるべきだが、生物学における定義では時間軸が厳格に規定されていないため、テリトリーという用語自体が曖昧さを持っている。行動圏(ホームレンジ)に関しても同様で、時間軸の定義が一週間なのか一年なのか、はたまた10年なのか、どこに設定して考えるかにじつは必然性が見出せない。生物学に文句をつけても仕方ないので、従来の生物学とは異なる時間が限られテリトリアルな空間が生じる場合に、その空間を「準テリトリー」と呼んで私自身はヒグマの空間利用の体系理解を進めてきた。
もうひとつ、テリトリーの拡張理論として種を越えた準テリトリー(もしくはテリトリー)を導入し、ヒトとヒグマの関係性について理解し共生モデルを考えてきた。
野生動物はそれぞれ様々な生存戦略を持ち、それを最大限に駆使して生き抜いてきたが、その性質や食物などによってテリトリーが合理的な場合と、不合理な場合とがある。テリトリアルな大型ほ乳類の代表はオオカミ、そうでない動物の代表はヒグマであるため私は対比によく使うが、ではヒトはどうなのか?と疑問が湧く。
通常、生物学的なテリトリーとは同種内に規定される概念だが、実際には、種を越えたテリトリーの存在が確認できるため、私はテリトリーの概念を種を越えたメカニズムとして拡張し考えている。
さて、問題のヒトであるが、現代のヒトは、特別な場合を除き間違いなくテリトリアルである。それも、かなり攻撃性の強い動物の一面を示している。例えば中山間地域で人里と呼ばれるのは農地を含んだ河岸段丘に沿ったヒトの活動範囲だが、そこに侵入してきたヒグマは侵入者として認識され、いきなり捕獲駆除という方法で排除にかかることが多い。比較的強いテリトリーを形成するオオカミでさえ進入個体を殺して排除などということはしない。ヒトが人里なりをヒトのテリトリーであると認識しているとしても、運用段階で根本的な不合理が現れる。つまり、テリトリーだと宣言するのはかまわないが、宣言だけではテリトリーは実質的に形成されない。宣言した範囲に関してオオカミがやるように誇示とメンテナンスをきっちりおこなわなければ絵に描いた餅でしかないのだ。ヒトが宣言したテリトリーがテリトリーとして機能するためには、オオカミのパトロール、ハウリング、マーキング、威圧的な追い払いなどが、ヒトにとっては何なのかを考え、電気柵・バッファスペース・追い払いやパトロールをおこなう必要がある。勝手に描いた絵の餅を巡って喧々諤々右往左往しているさまは滑稽ささえ漂わせている。 |
| |
あとがき
さて、ここに述べたヒグマの社会構造を決める忌避力学にヒトを混入させ統合して考える「忌避力学とテリトリーの拡張理論」は単なる理論かというと、じつは少なくとも2000~2017年の北大雪山塊・丸瀬布においてヒグマの生息地内において検証してきた理論であって、机上の空論ではない。アラスカ以来用いている「アクティブフェンス」と名付けた見えないフェンスを私自身は持っていて、それを自宅周辺に張ることによってヒグマの接近は確実に防げる。その確実さはアクティブフェンスの精度・強さによるが、同様にまわした電気柵と同等以上にその効果は発揮される。
電気柵より優れた点は、アクティブフェンスによる方法は自宅周りにヒグマが寄らなくなるだけでなく、私が山に入る場合にヒグマが自ずと私を避けて行動してくれることにある。電気柵はヒトに対する警戒心をヒグマの中に刷り込めないが、アクティブフェンス自体が私個人に対しての周辺ヒグマの忌避心理を用いているので、自宅にいても山に入っても安全に活動できるという結果が引き出せる。また、電気柵は単に電気柵に触れたクマが痛い思いをしてそれに触れることを避けるようになるだけで、逆に、触れなければ大丈夫というところまで学習させる。つまり学習能力のみを用いた方法だ。それに対して、アクティブフェンスはヒグマの高い類推能力に働きかけて作るフェンスで、必ずしも電気の刺激やスプレーや威嚇弾で痛い思いをさせなくても十分機能する。電気柵は受動的で、アクティヴフェンスは能動的である。そこで、アクティブフェンス(能動柵)と呼ぶことにした。アクティブフェンスはヒトそのものがつくることは困難だが、常にヒトと共にある文明によって比較的容易に作ることができる。
私の自宅を中心に半径5㎞の空間には、年間に25~30頭ほどのヒグマが侵入し活動している。入ったり出たりするので常時それだけの数がいるわけではないが、ヒグマの生息地の真っ只中と表現しても間違っていないだろう。ここに小屋を建て住み始めた頃は、100m以内にたびたびヒグマが現れた。そもそも、ここにはヒグマの好い場で、私がここに入る20年前からよく臥所も見られた場所。主要な移動ルートのひとつでもあった。周辺のクマにしてみれば、自分のお気に入りの寝床や通り道に怪しいヒトが独り居座りコツコツ何かやり始めたことが不服だったかも知れないし、微妙に興味をくすぐった部分もあったのかも知れない。しかし、数年後、ヒグマたちはここを迂回して移動するようになり、菜園に植えたスイートコーンやメロンなども荒らされたためしがない。そして現在、私自身は少し淋しいが400m以内にヒグマが接近することは皆無となっている。山にも集落方面にもクマは比較的多く活動し、集落方面ではクマ騒動や農地の被害が恒常化しているが、私の自宅を中心にポッカリとクマ不在の空間ができあがったことになる。膨大な観光客が訪れクルマが行き交う集落方面にクマは現れ、被害も及ぼす。その状況で、山側にクマ不在の空間が忽然とできあがる現象は、その原理が非常に重要だ。
科学的でないと言えば現段階ではその通りかも知れないが、自分がここに暮らすイメージとしては奥山に暮らす大型のオス成獣がある。つまり、メスや若グマが比較的高密度で暮らすこのエリアだが、そのクマたちに私が大型オス成獣のように思われていれば、それらは私から一定以上に距離をとって暮らすようになる。つまり、自らをヒグマの忌避力学や社会に混入させた形で暮らしているわけだ。
|
12+240+(18+780+18)+12=1080 780=28+724+28
|
|
| + |
| Copyright (C) 2014 higumajuku. All Rights Reserved. |
 |
| site map プライバシーポリシー 特定商取引法に基づく通信販売業者の表示――― powerd by higumajuku |

